日报标题:生殖隔离先不管了,我们来愉快地交配吧
 郭昊天,合成生物学博士在读/科学哲学/无伴奏合唱
郭昊天,合成生物学博士在读/科学哲学/无伴奏合唱
1. 生命演化史中有没有“生殖隔离”出现之后又消失的情况?
有,有很多。
在植物界生殖隔离出现后又被打破的例子实在是太多了(一会儿说为什么),但是我实在不喜欢素段子,所以还是举动物界的例子吧:
先写这个实验证据最明确的,存在的生殖隔离最清晰,也是最暴力的杂交例子:
1.1 北冰洋鸥属的进化与混乱杂交史
这个例子里面,生殖隔离出现后又被打破的是一群各种各样的鸥,其中的代表例子是银鸥和美国银鸥。
如果你知道环形种这个概念,一定对银鸥的例子不陌生:

这个表情严肃的鸟,其所在的鸥属有着非常混乱的杂交圈,他们分布在北冰洋周边,形成所谓的环形种(ring species):凡是地域相邻的两个群体之间都可以杂交,而不相邻的则不能杂交,而且有一对称为“终点”的不能杂交的这么一系列群体 / 物种。鸥属的其中一个环是这样的(摘自 Wikipedia):
1: 小黑背鸥 L. fuscus, 2: 也是小黑背鸥(西伯利亚分部)Siberian population of Larus fuscus, 3: L. heuglini, 4: 织女银鸥的一个亚种 L. vegae birulai, 5: 织女银鸥 L. vegae, 6: 美国银鸥 L. smithsonianus, 7: 银鸥 L. argentatus.
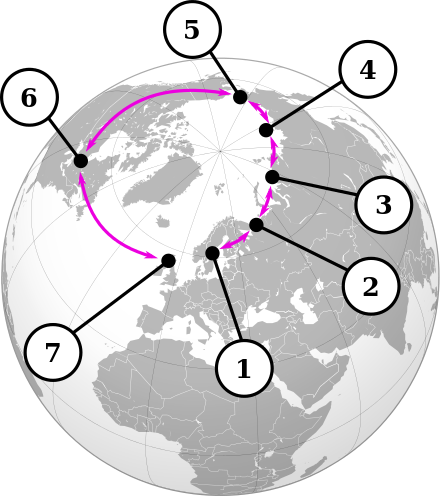
简单地讲,1 和 2 可以杂交,2 和 3 可以杂交,但是 1 和 3 因为空间隔离不会杂交,1 和 4 就更不可能了,到最后,小黑背鸥(1)和银鸥(7)的差异变得如此之大,以至于虽然生活在一个地方,但是完全不能互相杂交。我的 p 友的 p 友的……的 p 友,不是我的 p 友。
这种现象是怎样进化出来的呢?最简单的假说,就是认为啪啪的关系代表了进化的亲缘关系,可能发源地在欧洲,1 的一部分群体后来扩散到了西伯利亚演化成 2,2 的一部分群体又扩散到东西伯利亚演化成 3,……最后美国银鸥(6)中的一小撮飞回欧洲逐渐演化成银鸥(7)的时候,数万年前是一家的那些旧亲戚,已经不能一起愉快地大和谐了。这种假说也被称为环形种模型(ring-species model),演化过程中,随着迁移距离的增加,生殖隔离不断扩大,因此就可以很好解释 1 和 7 为什么不能杂交。早期假说基本上是这样一个思路。
但是!这些鸥其实并不符合“环形种模型”的解释。银鸥(7)不是美国银鸥(6)迁徙到欧洲后产生的“移民二代”,而是欧洲土著(Dorit Liebers et al 2004)。通过对大量样本做 DNA 分析,我们可以判定物种过去基因交流的模式以及发生的时间(就是一张写着诸如“xx 年王二狗和李小花生下了赵铁栓”的总谱),从而推断出进化和迁徙的途径,如下图所示:
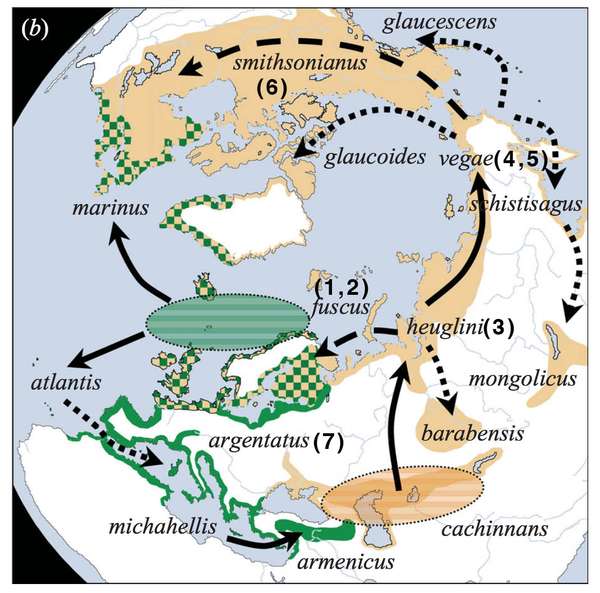
我们可以观察到进化的两个历程是 3-->1, 2-->atlantis-->7 和 3-->4,5-->6。也就是说,银鸥(7)和美国银鸥(6)是亲缘关系非常遥远的两个物种,他们的祖先之间有明显的生殖隔离,但是这并不妨碍他们现在能够一起愉快地生小鸥。这种杂交关系是后来重新建立的,由于地缘关系,可能是趋同进化导致的,但是具体原因并不清楚。不但如此,在北冰洋圈的这些鸥类之中,很多能够互相杂交的物种对,都不是进化关系上最近的亲缘物种。
由于各个鸥种之间还在发生着此消彼长的侵入,进化仍在进行,很有可能在不久的将来产生新的物种和杂交模式。
1.2 大中华区暗绿柳莺亚种演化史
Evolution and stability of ring species
1.3 鱼种形成大逆转,只因生活所迫http://www.nature.com/nature/journal/v482/n7385/full/nature10824.html
1.4 德州狼种乱杂交,也是生活所迫
Hybridization among Three Native North American Canis Species in a Region of Natural Sympatry
2. 生殖隔离是由基因组成决定的吗?
并不是。
什么是生殖隔离?就是要么两个群体之间不能互相交配,或者即使交配了后代也不能生育。生殖隔离的原因机制其实有非常多:
有行为差异是主要原因的,当然你可以通过一些手段逼良为娼,按住它们强行交配,很多也是能够获得可育后代。比如复旦的小白鼠突变体库里里有一个“屌丝突变”的公鼠,所有的母鼠都不愿意和它交配。它的身体其实是很好的,并没有精子不育、ED 这些毛病。如果人工授精,还可以获得同样是屌丝的“可育”后代。
有的是生理结构上的差异,比如 size 不匹配,你可以做人工授精,也是能够获得可育后代的。

这都属于交配前隔离。
还有一些属于交配后隔离,可以交配,但是要么不能形成受精卵,要么受精卵会变成死胎,要么生出来后代也不能生育。
基因组成对生殖隔离不是决定性的,有相同的基因也可能有生殖隔离,有不同的基因也可能没有生殖隔离。
毫无疑问,当基因组成差异非常大时,肯定是会有生殖隔离的。但是多大才算大呢?很多情况下,两个不能杂交的物种之间的基因差异,其实比他们种内的差异还小(比如上文的乱交鸥的例子)。在很多情况下,基因组成是不能决定最初的生殖隔离是怎样形成的。如果限定在基因决定的生殖隔离下讨论,就没的玩了……
相同组成的却又生殖隔离的典型例子就是染色体倍数变异。一旦亲本的精卵子提供的染色体不能配成对就会发生生殖隔离,而一旦能够配成对,就会恢复杂交的能力。所以,只要染色体数目一会儿减一倍,一会儿翻一倍,自然就可以产生“生殖隔离出现之后又消失”的情况了。由于植物发生染色体数目变异非常常见,所有很多这样的例子,比如我们日常吃的六倍体小麦。而在动物中,这种例子就很罕见了,毕竟植物的精子或者卵子掉地上可能长成一个新的植株,而我们就没这个能力了,掉地上只能默默地擦了。
3. 进化出相同的基因有多难?
因为题设是有问题的,所以高票的这个答案完全是被带坑里了。不过关于进化的一些设想和结论,也有错误。篇幅所限,我就不夸奖那些有价值的思考了,仅把错误的结论列出来。
3.1 进化并不是一条线
如果把每个基因的变化方式视为一个变量,进化就是在这个维度上亿的空间内一条不断转折的发散线。
首先要明白,进化的基本单位是群体,而不是个体。
进化过程中改变的是整个群体所有个体拥有的基因的“库”。
所以随机行走是一个非常糟糕的类比。一个无家可归的醉汉如果从酒吧出发,可能要花一晚上才能再次回到酒吧。但是一大群醉汉呢?总有那么一两个提前回到酒吧的。如果你养一大瓶细菌,不用多,10 毫升,所有可能的 DNA 单点突变,都会出现在这一瓶菌里。
因此绝不是一条线,而是”一群线”,两者的差别在于,当我们讨论他们的演化特征的时候,如果一个个体从 A 变成 B,另一个个体从 B 变成 A,从整个群体的角度,我们认为没有发生任何变化,但是如果认为每个个体都是独一无二的,则会给出不同的答案。(所以后面开的庞加莱重现时间的脑洞是不适用的)
3.2 选择!选择!选择!
基因“空间”的“体积”远远大于现有宇宙空间,两条线一旦分岔,几乎没有转回来的可能……这就是为什么基因变异可以用来测量物种分化的时间——大家默认基因一旦分化,就以恒定的速度在基因树上散开(抱歉文末提供了反例)。所以,两种动物通过随机变异消除生殖隔离……实现奇迹的概率比中彩票还低得多。
进化不是光有变异就完了的,达尔文心心念念的是自然选择啊!进化不是完全随机的,而是有选择方向的。
还是那个酒吧,还是那群醉汉。如果酒吧外面釜山行,那么所有活下来的醉汉都一定在酒吧里。
基因突变用来测量物种进化上的亲缘关系和分化时间,测量的往往都是快速进化的 DNA。这些 DNA 在序列结构上没那么保守,因此突变是相对中性的(不引起功能的丧失或改变),在连续地不断地突变,受选择影响小可以当做有相对恒定的突变速率。
但是如果这个 DNA 没那么重要,在不断地突变,很可能就没有一个质变的转捩点。如果没有这样一个分叉点,又怎么可能和生殖隔离有关系呢?
决定我是我,你是你的那些突变,一定都不会是中性的,一般都受到自然的强烈选择。而这些突变所在的基因,也不会是恒速进化的。比如我们和黑猩猩之间的唯一完全不同的基因FoxP2,决定我们有复杂的语言交流,而黑猩猩则没有这样的能力,使用FoxP2去估计我们和黑猩猩的分化时间是不能得到准确结果的。
这种受选择的突变,在特定的环境选择下,发生趋同进化,回复到同一点上,是很常见的。
其实在人类进化历程中就有很多这样的例子。
我在泥蚶红色的是血吗?的回答中还举过这样一个具体的例子:扁卷螺因为环境变动较大需要面对氧气不足的情况,在祖先已经丧失血红蛋白的情况下,重新进化出了血红蛋白。
3.3 趋同进化可以消除种间差异
1.3 中讲的就是这样一个例子。水华导致已经分化成不同物种的鱼最后融合成一种鱼了。
的确,理论上有很多不同的基因组合能够解决相同的问题,但是在现实的进化过程中要考虑初始的状态啊。
环境越极端(1.3 的例子里是水华污染),能够活下来的基因组合就越少,偏离这些组合死的越快(选择压强度远远超过了突变能产生的进化动力)。在这种情况下进化的结果就是陷入局部的最优组合。如果所有“物种”有着相同的共同祖先,能够触手可及的最优解都是一样的,那么他们毫无疑问最终会变成同一个“物种”。(P.S.这里物种不一定是真实的物种,也可能只是一大组基因序列组合)
在环境极端的时候有必然性,在环境不极端的时候就会有偶然发生的趋同消除种间差异的情况。因为进化的单位是群体,而且有选择发生,有优势的突变一旦在群体中出现要么还没传宗接代就车祸死了,要么就会逐渐扩散开来。因此这种偶然事件的出现概率也没有答主想像的那么低。
3.4 趋同进化有可能是普遍的,也有可能是强烈而必然的
但我依然认为,这是很偶然的现象,而且只是很小幅度的基因趋同。
进化过程中的事件的发生频率是几乎不可能准确报道的。
事实上,如果存在强烈的基因趋同效应,我们观察到的实验现象是:两个物种没有相邻的亲缘关系,生活在相似的环境中,拥有相同的基因。
而造成这个结果的机制,却并不是只有基因趋同一种,最简单的就是两个物种的相同基因来自共同祖先,只不过其他亲戚朋友们用不上这个基因所以把它丢掉了(后面将给出一个例子)。我们往往不能下结论是趋同进化的结果。
还有一些情况我们对基因的功能还不了解,连选择压力是什么都不知道,就更不可能判定趋同进化了。报道量较少的事情,也可能是广泛存在的,只不过限于目前的实验手段没有被广泛发现。
那么强烈而必然的趋同效应有没有呢?答案是肯定的,有很多情况下,只有一组合理的 DNA 序列,那么就没得选择,只能进化成这样。
一个简单的例子是启动基因表达的必需序列。不同基因的必需序列并不是来自相同的祖先,都是各自产生的,但是只要想表达基因,总得把序列变成这样,不管是从哪个祖先继承来的。一些以前没用的被沉默的假基因,通过这种突变就会恢复表达。我听到一个尚未发表的实验,通过使用人工合成的不能表达的基因进行定向进化,一两个礼拜就能产生新的表达,而新的表达,总是因为在某个地方产生了这组必需序列。
另一个例子是魏斯曼研究所 Dan S. Tawfik 组和 UW 的 David Baker 组的关于生命起源的研究成果(一时找不到是哪篇文章了,知道的朋友请评论区写一下,ty)。生命最开始的时候,DNA、RNA、蛋白质(多肽),他们要组装成一个生命系统,必须有相互作用。而这个最基本的核酸和蛋白质的相互作用的结构,可以通过从随机序列定向进化找到,也可以通过大量的计算进行搜索。最后的实验和计算分析结果证明,这个序列只有一个最优解(一个“关键序列”)。
这是个有趣的结果,但是让人头疼的问题也接踵而来。
我们对所以已知基因组的分析,说明在所有生物的共同祖先中也存在这个“关键序列”。那么我们现在看到的存在于各种各样的基因中的这段“关键序列”,到底是来自于共同祖先呢,还是从其他序列进化得来的呢。我们就不得而知了。
P.S. 关于@KittyFlier 提到的一个匿名回答中的看法,我需要澄清一下。首先动物间生殖隔离重新消失的原因我们并不完全清楚。趋同进化并不是充分的解释,在第 2 部分也提到了基因对于生殖隔离不是决定性的。我在最后这里只是对趋同进化的错误认知进行反驳而不是试图解释为什么恢复杂交能力。趋同进化导致产生相同的基因,甚至物种融合,在理论上没有那么困难。实际条件当然还是苛刻的,光靠趋同进化也不能完全解释生殖隔离消失,而几乎所有的例子只发生有一定亲缘关系的物种之间。
生态竞争中的动态变化是远远快于物种进化的,物种灭绝、共存等现象比恢复杂交要多 n 个数量级。除了灭绝之外的生态现象我们没有一个完全搞明白的。所以我们的讨论基础只能是,不考虑这些现象,如果这些物种的后代都活下来了,会是什么情景,是“理想条件”,不然就没得聊了╮(╯_╰)╭。